
Tempted by the serpent, Adam and Eve’s disobedience to God’s command not to eat from the Tree of the Knowledge of Good and Evil resulted in the serpent’s. eternally slithering, man’s eternally toiling, and as written in the King James Version of Genesis, “Unto the woman he said, I will greatly multiply thy sorrow and thy conception; in sorrow thou shalt bring forth children...” The story gives childbirth pain a moral and cosmic origin.
Modern science has moved away from sacred explanations for the pain and danger of childbirth. The dominant evolutionary account has long been the “obstetrical dilemma”: a trade-off between walking upright (bipedalism), which favors a relatively narrow pelvis, and giving birth to large-brained infants. In this view, the human birth canal is narrow, curved, and difficult to navigate because it must accommodate a baby whose head is relatively large compared with the mother’s anatomy. Childbirth, then, is not merely painful or medically risky; it is often presented as the evolutionary price paid for becoming human.
But science, for all its quantification, remains a human endeavor. The questions scientists ask, the comparisons they choose, and the stories they tell can all be shaped by cultural assumptions. One of the most persistent is human exceptionalism: the tendency to treat Homo sapiens as biologically unusual compared with other species. That anthropocentric view cast the obstetrical dilemma as a uniquely human struggle, resulting in high rates of maternal and neonatal risk compared with other members of our mammalian family.
Mammals vary widely in how developed their young are at birth. Altricial species, such as dogs, often give birth to litters of relatively helpless offspring. Precocial species produce young that are more developed and mobile soon after birth. Humans sit awkwardly on this spectrum: human babies are neurologically and physically dependent, yet they are also large-brained and energetically costly. One common explanation has been that human infants are born before they become too large to pass through the birth canal.
A recent review suggests that an anthropocentric bias has led us astray and that, contrary to our entrenched view, humans are not truly exceptional in their birth risks. Difficult births occur across placental mammals.
Dystocia: difficult birth across mammals.
Dystocia is the term for general birth complications and prolonged labor, in which the mother requires chemical or surgical assistance during delivery, typically due to a failure of “expulsive forces,” i.e., irregular uterine contractions or mechanical blockages from incorrect fetal positioning, such as a breech delivery or “fetopelvic” disproportion when the baby is simply too large to gracefully exit the birth canal.
The review surveys dystocia across mammalian species. It is limited because obstetric and veterinary medicine do not always classify the underlying cause of dystocia consistently. Mammalian births in the wild are difficult to document, so recorded cases are likely underreported. Human data are also compromised. Cesarean rates, influenced by local practice, liability concerns, and clinical judgment, are imperfect proxies. As a result, the review rightly treats the available data as cautious, broad comparisons.
What the mammal data show
The review finds that dystocia is a widespread mammalian problem; we are not alone. One might expect natural selection to eliminate a condition that can kill both mother and child. However, dystocia has been observed across a range of primates, as well as in deer and elephants. For some species, dystocia accounts for a substantial share of adult female deaths. Its persistence suggests that a difficult birth is not adaptive in itself but a recurring cost of reproductive strategies that are usually advantageous.
- In litter-bearing mammals, dystocia may arise from the trade-off between litter and offspring size. Small litters can produce larger individual offspring, increasing the risk of fetopelvic disproportion. Large litters can crowd the uterus, increase malposition, or contribute to uterine inertia. Precocial species give birth to more developed offspring that are larger, more mobile, and physiologically mature at birth, raising the mechanical demands of birth. A single large calf or neonate must pass through maternal anatomy that cannot expand without limit. In such species, fetopelvic disproportion and malpresentation become recurrent hazards.
- Dystocia has been documented in whales and other mammals whose pelvic anatomy differs markedly from that of humans. These animals have a reduced bony pelvis, and their birth problems cannot be explained simply by “pelvic constraint.” Instead, obstruction can also arise from soft tissues, fetal shape, fetal position, and the organization of the reproductive tract.
Biology sets the range of vulnerability, but inheritance, environment, and culture often determine where within that range a particular birth falls.
- Inheritance also shapes dystocia risk. This is evident in the selective breeding of domesticated mammals, which favors large offspring, rapid growth, unusual body shapes, or shortened skulls. Brachycephalic dogs, e.g., French bulldogs, experience dystocia at 16 times the rate of other dog breeds and have a 3-fold higher cesarean section rate because fetal head shape and maternal anatomy are poorly matched. [1]
- Environmental factors, including childhood nutrition, disease burden, obesity, and gestational diabetes, can shift the balance between fetal size and maternal capacity. Poor childhood nutrition may stunt growth and reduce pelvic dimensions, whereas nutritional excess during pregnancy can increase fetal growth. The most dangerous combination may be early undernutrition followed by adult overnutrition: a smaller maternal body paired with a larger fetus.
The Cliff-Edge Model of Selection
Difficult births are not confined to humans; they occur widely across mammalian orders, even when natural selection might be expected to have eliminated such a costly reproductive failure. The review reframes the challenges of birth as a broader, less anthropocentric question.
The “cliff-edge” model offers one explanation. Larger offspring often have better survival prospects, so selection can favor fetal growth. But birth imposes a hard limit: the fetus must still pass through the mother’s reproductive tract. Selection may 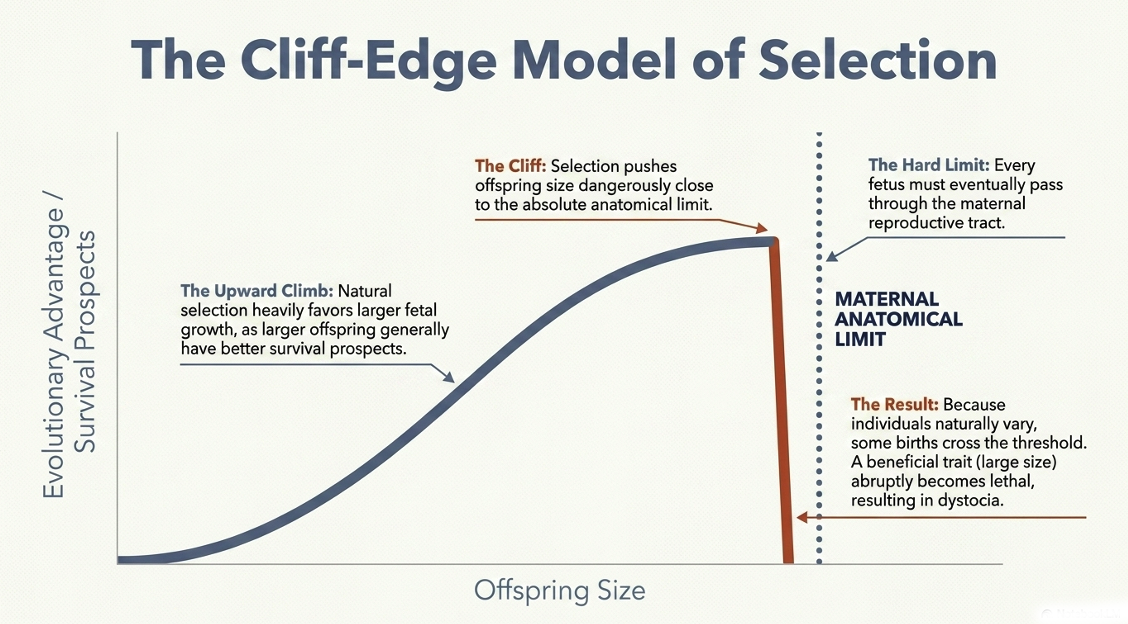 therefore push offspring size close to the edge of what maternal anatomy can tolerate. Because mothers and fetuses vary, some births inevitably cross that threshold. At that point, a trait that is usually beneficial, larger offspring size, becomes dangerous, and dystocia occurs.
therefore push offspring size close to the edge of what maternal anatomy can tolerate. Because mothers and fetuses vary, some births inevitably cross that threshold. At that point, a trait that is usually beneficial, larger offspring size, becomes dangerous, and dystocia occurs.
This model does not require human exceptionalism, nor does it require bipedalism or a large brain alone. Birth complications are an emergent property of placental reproduction, reflecting a persistent trade-off between the benefits of large offspring and the risks of a constrained birth canal.
Humans do face an “obstetrical dilemma,” but so do many other mammals. Difficult birth is not the strange exception that made us human; it is one expression of a much older mammalian compromise between the benefits of producing viable offspring and the dangers of bringing them into the world. Furthermore, each species’ environment modulates its baseline risk. In our case, the risk is intensified by malnutrition, obesity, or rapid nutritional transition, while it is reduced by midwifery, surgery, and medical care. What truly sets humans apart may not be the existence of difficult birth, but the scale of our biocultural response to it.
[1] “On average, 25% of puppies and 1.7% of the bitches do not survive problem births… with 48.6% of problem births needing caesarean surgery.”
Source: Humans are not unique: difficult birth is common in placental mammals Biological Reviews DOI: 10.1002/brv.70174

Chuck Dinerstein, MD, MBA
Director of Medicine
Dr. Charles Dinerstein, M.D., MBA, FACS is Director of Medicine at the American Council on Science and Health. He has over 25 years of experience as a vascular surgeon.


